


Forkhead box protein P2 (FOXP2) är en transkriptionsfaktor och tillhör gruppen forkhead box proteiner. FOXP2 upptäcktes först år 1998 vid studier av en Londonfamilj med många medlemmar, som led av svåra språkstörningar. Det är nu känt att FOXP2 spelar en viktig roll vid språkutveckling, däribland grammatisk förmåga.
FOXP2-genen kallas populärt för ”språkgenen”, men det skall inte övertolkas som att FOXP2 skulle vara ensam ”språkgen”. Många andra ryggradsdjur har också denna gen, och även hos dessa tycks FOXP2 spela en roll i verbal kommunikation. Avstängning av genen, till exempel hos möss - dessa kallas knockoutmöss - eller genom mutation hos människa, har en pleiotropisk effekt. Detta innebär att flera fenotypiska egenskaper ändrar sig. Vid sidan av hjärnan är FOXP2 också inblandad i utvecklingen av andra vävnader såsom lunga och inälvor.
Uppbyggnad
FOXP2-genen kodar för FOXP2-proteinet (se genetiska koden).
FOXP2-genen
Det uppskattas för närvarande att FOXP2-genen upptar minst 627 000 baspar (627 kb) på kromosom 7. Andra publikationer från 2007 indikerar antalet baspar till 603 kb. Oavsett vilket, utgörs ett stort antal baspar av introner, d.v.s. för proteinsyntesen funktionslösa genetiska element. Sjutton exoner, d.v.s. för proteinet kodande områden, återfinns.
Andelen 2145 (= 715×3) icke-kodande baspar i förhållande till det totalt ca 280 000 kodande baspar är med 0,6 % mycket låg men inte exceptionellt. Generellt varierar förhållandet mellan introner och exoner från gen till gen kraftigt. Så det finns vissa gener utan introner, medan andra består av mer än 95 % av introner.
FOXP2-proteinet
Det av FOXP2-genen kodade FOXP2-proteinet består av 715 aminosyror. Det är uppdelat i fyra huvudområden:
- en polyglutaminrik region, som består av två angränsande polyglutaminregioner och kodas av repetitiva CAG- och CAA-sekvenser,
- en zinkfinger-domän,
- en bZIP-domän (leucin-zipper) och
- en forkhead-domän som bildas av aminosyrorna 508 till 584.
Forkhead-domänen binds till DNA. Zinkfinger- och bZIP-domänerna är viktiga för protein-protein interaktioner och deltar också i DNA-bindningen.
Funktion
Som transkriptionsfaktor reglerar FOXP2-proteinet uppskattningsvis upp till 1000 andra gener, vilka är dock fortfarande till stor del okänt. Om konsekvenserna av ett skadat FOXP2 finns dock en gedigen kunskap.
Redan hos embryot återfinns FOXP2-proteinet. Det är främst uttryckt inom områden, som senare utvecklas till lillhjärnan (cerebellum), talamus (thalamus) och basala ganglierna (nuclei basales). Lillhjärnan och de basala ganglierna spela en viktig roll i inlärningen av komplexa motoriska färdigheter. För att kunna tala måste människan mödosamt lära sig en komplex motorik.
Tal- och språkutveckling
FOXP2-genen har i utvecklingen av språk- och talförmåga en central funktion. Därför medför mutationer i genen och en därav följande förlust av funktion hos proteinet till en specifik språk- och talstörning hos människan, särskilt inom artikuleringsförmåga och språkförståelse. Ett antal kända språk- och talstörningar, såsom autism, är därför kartlagda till FOXP2-genen på kromosom 7.
Schizofreni
Språkstörning är ett av de viktigaste symptomen på schizofreni. Av denna anledning och omedelbart efter upptäckten av FOXP2-genen misstänkte man att denna gen skulle kunna spela en viss roll för bildandet av schizofrenikänslighet. I en jämförande studie undersöktes 186 patienter med schizofreni (enligt Diagnostic and Statistical Manual of Mental Disorders DSM-IV hörde de konstiga röster) och 160 friska försökspersoner. Därvid analyserades speciellt FOXP2-genens nukleotid polymorfism. Statistiskt signifikanta skillnader angående frekvenser av genotyp (P = 0,007) och allel (P = 0,0027) mellan schizofrena patienter med hörselhallucinationer och kontrollgruppen kunde hittas i singel nukleotid polymorfismen (SNF) rs2396753. Resultatet tyder på att FOXP2-genen kan ha en inverkan på bildandet av schizofreni.
Upptäckt


År 1990 beskriver brittiska genetiker från Institute of Child Health i London en genetisk språkstörning, som drabbat tre generationer av en familj. Ungefär hälften av de 30 familjemedlemmarna hade stora problem med grammatik, syntax och ordförråd. I den vetenskapliga litteraturen kallas denna grupp för KE family (familjen KE). Denna bor i södra London och har pakistanskt ursprung. Den brittiske genetikern Anthony Monaco från University of Oxford i UK tillsammans med sin arbetsgrupp upptäckte år 1998 hos de familjemedlemmar, som hade språkstörningar, ett avsnitt av kromosom 7, som han knöt till familjens språkproblem. Genom genetiska studier av familjen KE och en ung pojke (patient CS), som inte har något samband med familjen KE men uppvisade samma symptom, kunde den så kallade ”språkgenen” FOXP2 identifieras för första gången.
Mutationen i genen FOXP2 uppträdde tydligt först hos mormor i familjen KE. Hennes språkstörningar var så allvarliga att även hennes man endast med svårighet kunde tyda hennes meningar. Alla tre döttrarna och en av hennes två söner har också språksvårigheter. Av de 24 barnbarnen visar tio samma symptom. De övriga medlemmarna i familjen från södra London har inga kommunikationsproblem. Den störning, som medlemmar i familjen KE uppvisar, kallas verbal utvecklingsdyspraxi (engelska: Developmental Verbal Dyspraxia (DVD)) och är införd i International Statistical Classification of Diseases and Related Health Problems som ICD-10 code F83 Mixed specific developmental disorders. En passande beskrivning på svenska är oförmåga att artikulera tal.
Symptom på en FOXP2-mutation
Verbal utvecklingsdyspraxi
Den allmänna beteendefenotypen hos människor med verbal utvecklingsdyspraxi visar sig i enkla tester med ord och ordupprepning. Därvid ska dikterade ord och icke-ord upprepas. Testpersonerna, som har mutationen, har betydligt större problem med sin artikulation än de som inte har mutationen. Försämringen ökar gradvis med komplexiteten i ordens artikulation.
Orofacial dyspraxi
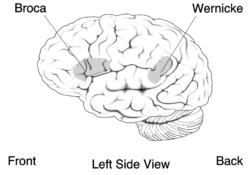
De berörda personerna har också svårigheter att styrt kontrollera ansiktsmusklerna, detta symptom kallas orofacial dyspraxi. Dessa svårigheter kan inte tillskrivas en allmän försämring av motorisk funktion, eftersom motoriska prestanda hos extremiteterna hos personerna inte går att skilja från normala individer. Även hörselförmågan hos patienterna är normal. DVD liknar den fenotyp, som man ser hos patienter med Brocas afasi. Det finns dock mellan de båda patologierna viktiga beteendemässiga skillnader. Personer med afasi är så mycket bättre i ordupprepningstestet än icke-ord upprepningtestet. De berörda medlemmarna med FOXP2-mutation i familjen KE är dock lika dåliga i båda typerna av test. En möjlig förklaring är, att människor med afasi har lärt sig i tiden före sin sjukdom kopplingen mellan mönster för ljudbildning och motsvarande ordbetydelser. Däremot har de drabbade medlemmarna i familjen KE hade aldrig möjlighet att lära mönster för artikulation av ord. Därför misslyckas de oundvikligen med att lösa de uppgifter med ordbetydelser i uppgifterna i ordupprepningstestet.
Andra symptom
Förutom verbal och orofacial dyspraxi är de av mutationen drabbade medlemmarna i familjen KE i tester, som utvärderingar de receptiva färdigheterna (språkförståelse) och färdigheter i att bilda grammatiska meningar, betydligt sämre än sina opåverkade släktingar. Bristerna inkluderar oförmåga att böja ord ordentligt, eller att bilda meningar, att fastställa enkla relationer mellan objekt och dess avbildningar. Dessutom uppvisar de berörda människorna i icke-verbal intelligenstest en betydligt lägre intelligens (intelligenskvot (IQ) genomsnitt: 86, intervall: 71-111) än de normala individerna (genomsnittlig IQ: 104, intervall: 84 till 119). Det finns en stor överlappning mellan de två grupperna.
Effekter av förändringar i FOXP2


De störningar som orsakas av mutationen ärvs autosomalt dominant. FOXP2-genen är belägen på den långa armen (q-armen) av kromosom 7 i band 7q31. Genen blev ursprungligen, eftersom endast den drabbade kromosomen kunde identifieras, kallad SPCH1 (speech-and-language-disorder-1).
Vid en år 2006 genomförd screening av 49 försökspersoner, som led av verbal dyspraxi, hittades en försöksperson med en på mödernesidan ärvd nonsense mutation och två försökspersoner med förment missense mutation av FOXP2-genen. Dessa resultat tyder på att FOXP2 mutationer är en relativt ovanlig orsak till tal- och språkstörningar.
Exon 14 (familjen KE)
De genom arv sjuka medlemmarna av familjen KE har en missense punktmutation i exon 14 av genen. Nukleinbasen guanin ersätts hos dem på en plats med adenin. Därigenom blir på plats 553 av FOXP2-proteinet aminosyran histidin installerad i stället för aminosyran arginin. I aminosyrors en-bokstavs-förkortning är namnet på arginin R och på histidin H. Mutationen har därför fått namnet R553H. Det bildade proteinet kan till följd av bytet av aminosyra inte längre fylla sin funktion.
Vid avbildning av hjärnan hos medlemmar av familjen KE kunde avvikelser observeras i svanskärnan (nucleus caudatus), en del av de basala ganglierna. De första insikterna i de neurala grunderna kunde fås genom funktionell magnetresonanstomografi (fMRT) (engelska: functional Magnetic Resonance Imaging (fMRI)) av hjärnan. De av mutationen berörda medlemmarna av familjen KE uppvisar bilaterala strukturella brister. Detta visade sig främst i en minskad täthet av grå hjärnsubstans (substantia grisea) i svanskärnan i basala ganglierna, i främre delen av lillhjärnan (cerebellum) och i Brocas område. Däremot hittades hos dessa patienter en onormalt hög täthet av grå hjärnsubstans i skalkärnan (putamen) och i Wernickes område. Intressant är att volymen av svanskärnan korrelerar mycket väl med i språktest visad förmåga. Detta är en indikation på svanskärnans inflytande på den verbala utvecklingsdyspraxins (DVD) patologi.
De basala ganglierna spela en avgörande roll i planering och sekvensering av rörelse. Strukturella variationer i striatum-regionen i de basala ganglierna (svanskärnan och skalkärnan) innebär därför i allmänhet en effekt på kontrollen av orofacial motorik (munmotoriken). Det är dock oklart varför specifikt munmotoriken är nedsatt, utan att andra motoriska funktioner påverkas.
Exon 7 (nonsense mutation)
År 2005 upptäcktes hos barn som inte hör till familjen KE, men likväl lider av en verbal dyspraxi, en så kallad nonsense mutation också i FOXP2-genen. En nonsense mutation är en förvrängande mutation, en stoppkodon bildas, följaktligen en bas-trippel, vilket leder till stopp av syntesen av protein vid denna plats. Även i dessa fall med nonsense mutation hänförs tal- och språkstörningarna till mutationen.
Brytningen mellan exon 3b och 4
En patient, känd i litteraturen som patient CS, har en balanserad translokation av kromosomerna 5 och 7 (t(5;7) (q22;q31.2)). Brytpunkten på kromosom 7 är i FOXP2-genen mellan exon 3b och 4 och påverkar därmed alla kända isoformer av FOXP2-proteinet. Denna patient lider också av liknande symptom som de av mutation drabbade medlemmar i familjen KE.
Gendeletion
Förlusten av en bit DNA i en kromosom kallasdeletion. Hos en kanadensisk flicka visade sig år 2006 ha en förlust (deletion) av avsnitt av kromosom 7 i band 7q31 och 7q32. I det felande området återfinns också FOXP2-genen. Barnet har svåra kommunikationsstörningar i form av en orofacial dyspraxi, en betydande vanskapnad och en försening i sin allmänna utveckling. Hon kan inte hosta, nysa eller skratta spontant.
Hos möss

Hos möss avstängdes både exon 12 och 13 i FOXP2 (knockout) i djurförsök. Om båda kopiorna av FOXP2-genen avstängdes, ledde detta till allvarliga störningar i motorik, tidig död orsakad av otillräcklig lungutveckling och brist på kommunikation med ultraljud. Det senare kan oftast ge upphov till att unga djur överges av sin mamma. Var dock bara en av de båda kopiorna av FOXP2-genen avstängd, ledde detta till en ringa fördröjning i utvecklingen av djuret och en betydande förändring i kommunikationen med ultraljud. Djuren har onormala förändringar i lillhjärnan (cerebellum), särskilt purkinjecellerna (gangliecellerna i Stratum gangliosum i Gyri cerebellares).
Vid inplantering av den mänskliga varianten av gener i genomet hos möss visade dessa en signifikant förbättring i inlärningsförmåga jämfört med oförändrade djur. Därvid kunde ses förändringar i de basala ganglierna hos de förändrade mössen.
Hos zebrafinkar

Språkutveckling är inte begränsat till människor. Vissa djurarter, däribland valar, småfladdermöss och fåglar, från tre biologiska ordningar kan lära sig sin akustiska kommunikation ("djurspråk") genom imitation.Sångfåglar kommunicerar genom sång som de i stor utsträckning måste lära sig. De förvärvar sina ljudsekvenser genom att de imiterar äldre artfränder. Unga fåglar, som är isolerade från artfränder, blir följaktligen stumma. Sångfåglar är därför lämpliga som djurmodell (modellorganism) för studier av språkutveckling och dess genetiska anlag. Hos många andra arter är dock läten medfödda. Även hos apor antas det, att deras repertoar av ljud är medfödd.
Zebrafinkens (Taeniopygia guttata) sång består av olika stavelser, som resulterar i strukturerade sekvenser. Det viktiga hjärnområdet för språkutveckling hos människan finns i de basala ganglierna. Hos sångfåglar är detta område känt som Area X. Genuttrycket av FOXP2 i Area X är som högst under zebrafinkens sånginlärningsfasen. Hos kanariefåglar är genuttrycket av FOXP2 varierat säsongsvis. I perioder när sången ändras är genuttrycket särskilt högt. Hos fågelarter, som inte lär sig sin sång, såsom ringduvan, har inga jämförbara förändringar i FOXP2 genuttryck kunnat fastställas.
Med hjälp av RNA-interferens stängde forskare på Max Planck Institute for Molecular Genetics i Berlin av FOXP2-genen i Area X på zebrafinkar. I den här processen har korta kompletterande komplementära RNA segment införts i cellerna, där de fångar mRNA och undertrycker produktionen av FOXP2-protein. Zebrafinkarna, som hade avstängd FOXP2, imiterade stavelserna hos sina äldre artfränder med låg precision och utelämnade hela stavelser i sin sång.
Den exakta verkningsmekanismen hos FOXP2 är ännu inte känd. I princip kan den genetiska defekten påverka den motoriska funktionen, till exempel röstorganet, eller sparandet av sånger avsedda att läras.
Den molekylära evolutionen av FOXP2

FOXP2 hos däggdjur och andra ryggradsdjur
FOXP2-proteinet hos däggdjur tillhör till de väl bevarade (engelska: highly conserved) proteinerna. Det skiljer sig endast mycket obetydligt mellan olika arter. Ett undantag utgör olika familjer av fladdermöss. Mellan dem finns betydande skillnader i FOXP2-sekvensen. Å andra sidan kan man hitta nästan identiska FOXP2-proteiner till exempel hos sångfåglar, fiskar och kräldjur.
Polyglutamin är uppbyggd av längre återkommande glutamin-glutamin-länkar. Gensegment, som kodar för polyglutamin, är generellt kända för att de har relativt hög mutationsfrekvens. Detta är också fallet för de två polyglutamin-regionerna hos FOXP2-genen. Så alla studerade taxa visade olika polyglutamin-längder. För FOXP2-proteinets funktion spelar polyglutamin-regionen en mycket liten roll. Om man ignorerar dessa regioner skiljer sig det mänskliga FOXP2-proteinet från musens ortolog endast i tre aminosyror.
Den evolutionära linjen, som leder till människa och mus, delades för att omkring 40 miljoner år sedan. Den senaste gemensamma förfadern till schimpans och människa levde för 4,6 till 6,6 miljoner år sedan.Av de tre aminosyrornas skillnad mellan människa och mus finns en aminosyras skillnad till musens förfader, ingen skillnad mellan musens och primaternas förfader och delningen av människan och schimpansen och två aminosyrors skillnad därefter (se figuren till höger). FOXP2-proteinet hos orangutangen avviker med två aminosyror från musens och med tre från människans. Även i FOXP2-proteinet hos zebrafinkar finns bara sju aminosyrors skillnad jämfört med människans.
Möjligheten för människor att kunna tala, är baserad på anatomiska och finmotoriska färdigheter, som andra primater vid sidan av människan inte har. Vissa forskargrupper antar att de två aminosyrornas skillnad mellan schimpans och människa har lett till språkutveckling hos människor. Denna hypotes är emellertid omdiskuterad eftersom andra arbetsgrupper inte fann någon koppling mellan arter med inlärda läten och de med liknande mutationer i FOXP2.
De två skillnaderna mellan människan och hennes närmaste släktingar finns i exon 7. På position 303 har treonin ersatts med asparagin och på position 325 asparagin med serin. De sannolika proteinstrukturerna bestämdes genom beräkningar genom simulering. Mutationen i position 325 producerar en potentiellt reaktiv plats för fosforylering av proteinkinas C i det mänskliga FOXP2-proteinet tillsammans med en liten förändring i sekundärstrukturen av proteinet. Från olika studier är det känt att fosforyleringen av transkriptionsfaktorer med forkhead-struktur kan vara en viktig mekanism i genregleringen. För att klargöra om de två aminosyror kodade i exon 7 hos människan är polymorfa har detta exon sekvenserats för 44 personer från olika kontinenter. I inget fall hittades någon form av aminosyra-polymorfism.
Fladdermöss


Medan hos de flesta däggdjur genom systematisk DNA-sekvensering kan en extremt låg - det handlar om endast ett fåtal aminosyror - mutationsfrekvens fastställas i FOXP2-genen, har betydande skillnader identifierats hos vissa arter av fladdermöss. Fladdermössen är bland de få ryggradsdjur som har förmågan att lära sig ljud.
Den biologiska ordningen av fladdermöss (Chiroptera) består av två underordningar: fruktfladdermöss eller flyghundar (Megachiroptera) och småfladdermöss (Microchiroptera). Småfladdermöss använder ekolokalisering till orientering och för att fånga byten. Hos dem är de sensomotoriska förmågorna särskilt väl utvecklade. Mottagning av överförda ultraljud kräver ett utmärkt hörselsinne och - beroende på småfladdermusart - orofacial (mun) eller nasofacial (näsa) samordning. Flyghundar har däremot ingen förmåga till ekolokalisering.
Vid DNA-sekvensering identifierades de båda exonerna 7 och 17 som områden i vilka - beroende på fladdermusart - den största variationen av FOXP2-genen fanns. Det fanns signifikanta skillnader mellan FOXP2-strukturen hos småfladdermöss och flyghundar. Data leder till slutsatsen att förändringarna av FOXP2-genen hos småfladdermössen har spelat en avgörande roll för utveckling av ekolokalisering.
FOXP2 i paleogenetiken
Paleogenetik är en disciplin som använder genetiska metoder för att studera tidiga Homo arter och andra forntida populationer. Med hjälp av paleogenetik beräknades först att genvarianten av FOXP2, som är vanlig idag hos människor, är mellan 100 000 och högst 200 000 år gammal. Denna period utforskades i en matematisk modell där specifikt mutationer i introner undersöktes. Introner är genetiska element, som inte har någon funktion i proteinsyntesen. Eftersom de inte har betydelse på strukturen av proteiner, kan en mycket högre mutationsfrekvens observeras hos dem än hos exoner. Från denna mutationsfrekvens kan en gens historia rekonstrueras. Den beräknade tidsperioden skulle väl sammanfalla med den av paleoantropologer daterade "födelsedagen" för människan som art (Homo sapiens). Detta är betydligt senare än tidpunkten, likaså bestämd genom paleogenetik, för det evolutionära stamträdets uppdelning mellan Homo sapiens och Homo neanderthalensis satt till mellan cirka 400 000 år upp till kanske 800 000 år sedan. Utifrån dessa uppgifter drogs först slutsatsen att neandertalarna inte hade människans språkförmåga.
Några antropologer hävdar att den snabba spridningen av den så grundläggande för språkutveckling FOXP2-genen talar för tesen att språket har varit den drivande kraften bakom spridandet av människan på jorden.
Tesen om åldern av moderna människans FOXP2 genvariant och av den dra slutsatsen att neandertalarna därför inte hade människans språkförmåga fick revideras i oktober 2007. FOXP2-genen har sekvenserats från neandertalares ben. Då hittades ingen skillnad i neandertalarens sekvens i jämförelse med den moderna människans. Uppkomsten av någon form av språk kan därför uppskattas till ca 400 000 år upp till kanske 800 000 år sedan.
DNA-sekvensering av förhistoriska fynd är en mycket komplicerad process. Proverna innehåller mycket små mängder endogen DNA. Dessutom är kontaminering av prover och reagenser med mänskligt DNA ett betydande problem, särskilt som att neandertal-DNA skiljer sig väldigt lite från den moderna människans. Två olika cirka 43 000 år gamla neandertalben, som år 2006 hittades i Sidrón-grottan (spanska: Cueva de El Sidrón) i Asturien i nordvästra Spanien, analyserades först avseende mitokondriellt DNA (mtDNA). Baserat på mtDNA kan det avgöras genom några välkända substitutioner (utbyten), om det är DNA från en modern människa eller en neandertalare. När det var klart, att proven tydligt innehöll neandertal-DNA, analyserades de två områden i exon 7 av FOXP2-genen, som är kända för mutationer efter separationen av människa och schimpans. Samtidigt hittades ingen skillnad i motsvarande sekvenser från neandertalare och modern människa. Neandertalare hade således också mutationen av FOXP2-genen, som medger språk. Möjligheten, att genen genom gemensamma barn till Homo sapiens och Homo neanderthalensis inblandats både hos moderna människor och hos neandertalare, är uteslutet på grundval av resultat från studier av mitokondriellt DNA.
Interaktioner
FOXP2 har visat sig interagera med protein CTBP1.
Litteratur
Fackartiklar
- G. Konopka et al.: Human-specific transcriptional regulation of CNS development genes by FOXP2, Nature, 462, 2009, sid. 213–217. PMID 19907493
- C. S. Lai et al.: A forkhead-domain gene is mutated in a severe speech and language disorder, Nature, 413/2001, sid. 519-23. PMID 11586359
- F. Vargha-Khadem et al.: FOXP2 and the Neuroanatomy of Speech and Language, Nature Reviews Neuroscience, 6/2005, sid. 131–138. PMID 15685218
- F. Vargha-Khadem et al.: Praxic and nonverbal cognitive deficits in a large family with a genetically transmitted speech and language disorder, Proc Nat Acad Sci, 92/1995, sid. 930-33. PMID 7846081
- H. A. Bruce, R. L. Margolis: FOXP2: novel exons, splice variants, and CAG repeat length stability, Human Genetics, 111/2002, sid. 136-44. PMID 12189486
- C. S. Lai et al.: A novel forkhead-domain gene is mutated in a severe speech and language disorder, Nature, 413/2001, sid. 519-23. PMID 11586359
- B. Wang et al.: Multiple domains define the expression and regulatory properties of Foxp1 forkhead transcriptional repressors, J Biol Chem, 278/2003, sid. 24259-68. PMID 12692134
- J. C. Stroud et al.: Structure of the forkhead domain of FOXP2 bound to DNA, Structure, 14/2006, sid. 159-66. PMID 16407075
Fackböcker
- W. Bigenzahn, G. Böhme: Sprach-, Sprech-, Stimm- und Schluckstörungen Elsevier Deutschland, ISBN 3-437-46950-9
- R. J. McCauley: Assessment of Language Disorders in Children, Lawrence Erlbaum Associates, 2001, sid. 118, ISBN 0-8058-2562-2
- Stefanie Otte: Gibt es Zusammenhänge zwischen einer expressiven Sprachentwicklungsstörung und einem zentro-temporalen Sharp-Wave-Fokus (Rolando-Fokus) mit der weiteren Entwicklung? Dissertation, Julius-Maximilians Universität zu Würzburg, 2005, sid. 82-83.
- Harald Teepe: Welche Bedeutung haben die Neurowissenschaften für die Fremdsprachendidaktik?, Dissertation, Rheinisch-Westfälische Technische Hochschule Aachen, 2005
- Sverker Johansson: Origins of Language - Constraints on hypotheses, Benjamins Publishing, 2005, ISBN 90-272-3893-6
Populärlitteraur
- Mason Inman: Neandertals Had Same "Language Gene" as Modern Humans, National Geographic News, 18 oktober 2007
- Bijal P. Trivedi: Scientists Identify a Language Gene, National Geographic News, 4 oktober 2001
- Lotta Fredholm: Vår utdöda släkting väcks till liv, Forskning och Framsteg, 5/2009
- Per Snaprud: Utan språkgen sluddrar finkarna, Forskning och Framsteg, 6/2008
- Lotta Fredholm: Neandertalarna kunde kanske tala, Forskning och Framsteg, 1/2008
- Annika Nilsson: Neandertalarna närmare oss än vi trott, DN.se, 2008-11-29
- Ulrika Kahl: Ännu för tidigt säga vilka »talgener« som styr människans språkförmåga, Läkartidningen, 12 december 2002
Externa länkar
- FOXP2 i RCSB Protein-Data Bank
- K. Seefeldt: Der kleine Unterschied Telepolis,16 augusti 2002
- A. MacAndrew: FOXP2 and the Evolution of Language
- Our Recent Research on FOXP2 vid University of Oxford
- Jane Itzhaki: The FOXP2 story Wellcome Trust Centre for Human Genetics, University of Oxford, 28 april 2003
- Revisiting FOXP2 and the origins of language
- Hemsida Max Planck Institute for Evolutionary Anthropology
- Den här artikeln är helt eller delvis baserad på material från tyskspråkiga Wikipedia, Forkhead-Box-Protein P2, 31 december 2011.